
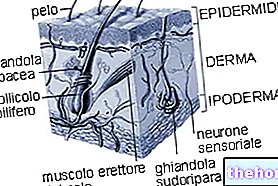
Näiden lieriömäisten yksiköiden ansiosta aineenvaihduntareaktioiden vapauttama kemiallinen energia muuttuu mekaaniseksi energiaksi; työnnettäessä itsensä jänteiden läpi ja vaikuttamalla luuvipuihin lihas tuottaa liikettä.
Luustolihasten pituudet vaihtelevat muutamasta millimetristä useisiin senttimetreihin ja niiden halkaisija on 10-100 µm (1 µm = 0,001 mm); ne ovat kehon suurimmat solut.
"Sytologisesti" kuitusolut ovat seurausta prosessista, jota kutsutaan myogeneesiksi, joka on useiden myoblastien fuusio - toiminta, joka riippuu lihaksispesifisistä proteiineista, joita kutsutaan fusogeenit, myomaker tai myomerger. Siksi myosolut näyttävät pitkiltä lieriömäisiltä ja monisoluisista soluista (jotka sisältävät lukuisia myonukleoita - muun muassa selvästi näkyvissä mikroskoopin pinnalla).
Lihaskuitu, esim. haaran hauisissa, joiden pituus on 10 cm, sillä voi olla jopa 3000 ydintä.
Niiden sisällä on sen sijaan tuhansia filamentteja, nimeltään myofibrillejä, jotka sisältävät supistuvia yksiköitä, joita kutsutaan sarkomeereiksi.
Lihaksia käsittelevät fysiologit kertovat meille, että eri kuidut eroavat toisistaan paitsi anatomisesti myös joidenkin tarkkojen fysiologisten ominaisuuksien vuoksi.
Siksi jokaisessa lihassa tunnistetaan erityyppisiä kuituja, jotka on luokiteltu eri kriteerien mukaan, kuten energian aineenvaihdunta, supistumisnopeus, väsymiskestävyys, väri jne.
Kaiken kaikkiaan yksi lihas, kuten esim. haaran hauis, noin 253 000 lihaskuitua.
Tiesitkö että ...
Tyvikalvon ja lihaskuitujen sarkolemman välissä on ryhmä lihaskantasoluja, jotka tunnetaan myosatelliittisoluina.
Nämä ovat yleensä lepotilassa, mutta ne voidaan aktivoida liikunnan tai sairauden avulla, jotta saadaan lisää lihassolujen kasvua tai korjaamista varten tarvittavia myonukleoja.
spesifiset fosfaagit (ATP ja CP), mitokondriot, myoglobiini, glykogeeni ja suurempi kapillaaritiheys.
Lihasolut eivät kuitenkaan voi jakautua uusien solujen tuottamiseksi, ja sen seurauksena niiden määrä pyrkii pienentymään iän myötä.
), jotka aiheuttavat kolme kuitutyypit.
Näillä kuiduilla on suhteellisen selviä aineenvaihdunta-, supistumis- ja moottoriominaisuuksia - yhteenveto alla olevasta taulukosta.
TÄRKEÄ! Eri ominaisuudet, vaikka ne riippuvatkin osittain yksittäisten kuitujen ominaisuuksista, ovat yleensä merkityksellisempiä moottoriyksikön tasolla mitattuna - mutta niissä on kuitenkin hyvin vähän vaihtelua kuitujen monimuotoisuuden suhteen - eikä yksittäinen kuitu.
Katsotaan nyt joitain luokittelutyyppejä.
Kuitu väri
Perinteisesti kuidut luokiteltiin niiden värin mukaan, mikä riippuu myoglobiinipitoisuudesta.
Tyypin I kuidut näyttävät punaisilta korkeiden myoglobiinitasojen vuoksi, niillä on yleensä enemmän mitokondrioita ja suurempi paikallinen kapillaaritiheys.
Ne kutistuvat hitaammin, mutta sopivat paremmin vastustuskykyyn, koska ne käyttävät hapettavaa aineenvaihduntaa ATP: n (adenosiinitrifosfaatin) tuottamiseksi glukoosista ja rasvahapoista.
Vähemmän hapettavat tyypin II kuidut ovat valkoisia tai joka tapauksessa kirkkaita myoglobiinin niukkuuden ja glykolyyttisten entsyymien pitoisuuden vuoksi.
Supistumisen nopeus
Kuidut voidaan luokitella supistumisnopeutensa mukaan nopeiksi ja hitaiksi. Nämä piirteet ovat suurelta osin, mutta eivät kokonaan, päällekkäisiä väriin, ATPaasiin ja MHC: hen perustuvien luokitusten kanssa.
- Kuidut a nopea supistuminen, jossa myosiini voi jakaa ATP: n hyvin nopeasti. Näitä ovat tyypin II ATPaasi ja tyypin II MHC -kuidut. Ne osoittavat myös suurempaa kapasiteettia toimintapotentiaalien sähkökemialliseen siirtoon ja nopeaa kalsiumin vapautumista ja imeytymistä sarkoplasmisen verkkokalvon kautta.Ne perustuvat hyvin kehitettyyn, anaerobiseen, nopeaan energiansiirtoon perustuvaan glykolyyttiseen järjestelmään ja voivat supistua 2-3 kertaa nopeammin kuin hitaat nykimiskuidut Nopeat lihakset sopivat lyhyiden voima- tai nopeusjaksojen tuottamiseen kuin hitaat lihakset ja siten väsymys nopeammin.
- Kuidut a hidas supistuminen tuottaa energiaa ATP: n uudelleensynteesille aerobisen ja pitkäkestoisen siirtojärjestelmän kautta. Näitä ovat pääasiassa tyypin I ATPaasi- ja tyypin I.Hitaasti nykivät kuidut kehittävät enemmän mitokondrioita ja kapillaareja, mikä tekee niistä parempia kestävyystyöhön.
Kuitujen kirjoitusmenetelmät
Kuitujen kirjoittamiseen käytetään useita menetelmiä, mikä usein aiheuttaa epäselvyyttä ei-asiantuntijoiden keskuudessa.
Kaksi usein epäselvää menetelmää ovat myosiini -ATPaasiaktiivisuuden histokemiallinen värjäys ja myosiinin raskaan ketjun tyypin (MHC) immunohistokemiallinen värjäys.
Myosiini -ATPaasientsyymin aktiivisuutta kutsutaan yleisesti ja oikein yksinkertaisesti "kuitutyypiksi", ja se johtuu ATPaasientsyymin aktiivisuuden suorasta mittaamisesta eri olosuhteissa (esim. PH).
Myosiinin raskaan ketjun värjäystä kutsutaan tarkemmin "MHC -tyypiksi" (myosiinin raskas ketju), ja kuten voidaan ymmärtää, se johtuu eri MHC -isoformien määrittämisestä.
Nämä menetelmät liittyvät fysiologisesti, koska MHC -tyyppi on ATPaasiaktiivisuuden tärkein määrittävä tekijä. Mikään näistä kirjoitusmenetelmistä ei kuitenkaan ole luonteeltaan suoraan metabolista; tuo on ne eivät suoraan käsittele kuidun oksidatiivista tai glykolyyttistä kapasiteettia.
Kun viitataan "tyypin I" tai "tyypin II" kuiduihin, tämä viittaa tarkemmin arviointiin värjäämällä "myosiinin ATPaasiaktiivisuus" (esim. "Tyypin II" kuidut viittaavat tyypin IIA + tyypin IIAX + tyypin IIXA ... jne.).
Alla on taulukko, joka esittää näiden kahden menetelmän välisen suhteen, rajoittuen ihmisissä oleviin kuitutyyppeihin. Jotkut ATPaasi -tyypit sisältävät itse asiassa useita MHC -tyyppejä.
Lisäksi alatyyppiä B tai b ei ekspressoida ihmisillä kummallakaan menetelmällä. Varhaiset tutkijat uskoivat, että ihmiset voivat ilmaista MHC IIb: tä, mikä johti IIB: n ATPaasiluokitukseen. Myöhemmät tutkimukset ovat kuitenkin osoittaneet, että ihmisen MHC IIb on itse asiassa IIx, mikä osoittaa, että oikeampi sanamuoto on IIx.
Alatyyppi IIb tai IIB, IIc ja IId ilmaistaan sen sijaan muissa nisäkkäissä, kuten kirjallisuudessa on laajalti dokumentoitu.
Muut kuitujen kirjoittamismenetelmät on hahmoteltu vähemmän muodollisella tavalla ja niitä on enemmän, kuten yleisurheilukentällä normaalisti käytetty.
Niillä on taipumus keskittyä enemmän aineenvaihdunta- ja toimintakykyihin (supistumisaika, pääasiassa hapettava vs. anaerobinen laktaatti vs. anaerobinen laktaatti, nopea vs. hidas supistumisaika).
Kuten edellä todettiin, kuidun kirjoittaminen ATPaasin tai MHC: n avulla ei suoraan mittaa tai määrää näitä parametreja. Kuitenkin monet eri menetelmistä liittyvät mekaanisesti, kun taas toiset liittyvät toisiinsa in vivo.
Esim, ATPaasikuidun tyyppi liittyy supistumisnopeuteen, koska ATPaasin korkea aktiivisuus mahdollistaa nopeamman poikkisillan kierron. Tyypin I kuidut ovat "hitaita" osittain siksi, että niillä on alhainen ATPaasiaktiivisuus verrattuna tyypin II kuituihin; supistumisnopeuden mittaaminen ei kuitenkaan ole sama asia kuin ATPaasikuidun kirjoittaminen.
, valkoisia ja välituotteita. Niiden osuudet vaihtelevat kuitenkin sen lihaksen fysiologisen työn mukaan.Esimerkiksi ihmisillä nelipäiset lihakset sisältävät noin 52% tyypin I kuituja, kun taas pohjapohja on noin 80%.
Tiesitkö että ...
Lihaskuidun kehittämä voima riippuu sen pituudesta supistumisen alussa. Sen on oltava optimaalinen arvo, jonka ulkopuolella (vetäytynyt tai liian venytetty lihaksen) voima on heikentynyt. Lihasten vahvistamisen alalla yleisin virhe on treenata lihaksia jo osittain lyhennettynä. Ainoat poikkeukset sääntöön ovat kivun tai epämukavuuden esiintyminen tai paramorfismit, jotka siksi vaativat liikerajan rajoittamista (ROM).
Pääasiassa valkoisia lihaksia, joissa on runsaasti tyypin II kuituja, kutsutaan vaiheiksi, koska ne kykenevät nopeisiin ja lyhyisiin supistuksiin. Toisaalta punaisia lihaksia, joissa tyypin I kuidut ovat vallitsevia, kutsutaan tonikoiviksi, koska ne kykenevät supistumaan pitkään.
Lihaksen moottoriyksiköissä on kuitenkin hyvin vähän vaihtelua, mikä tekee moottoriyksiköiden rekrytoinnin ulottuvuusperiaate; toisin sanoen keho kykenee tarvittavasta voimakkuudesta / voimakkuudesta riippuen stimuloimaan vain osaa (esim. pitkäaikaisesta aerobisesta toiminnasta) tai kaikkia (esim. suurimman kyykkyn aikana) kyseisiä yksiköitä.
Nykyään tiedämme, että kuitujen jakautumisessa ei ole sukupuoleen liittyviä eroja. Kuitenkin eri tyyppien suhteet - jotka tiedämme vaihtelevat suuresti eläinlajien ja vähemmässä määrin etnisten ryhmien välillä - "voivat" vaihdella huomattavasti henkilöstä toiseen.
Joidenkin oivallusten mukaan istuvilla miehillä ja naisilla (samoin kuin pienillä lapsilla) pitäisi olla 55% tyypin I kuitua ja 45% tyypin II kuitua.
Toisaalta korkean tason urheilijoilla on erityinen kuitujakauma käytetyn aineenvaihduntatyypin perusteella. Maastohiihtäjillä on pääasiassa kuitu I, pikajuoksijoilla pääasiassa II ja keskimatkan juoksijoilla, heittäjillä ja hyppääjillä, jotka ovat lähes päällekkäisiä.
Siksi on ehdotettu, että erilaiset harjoitustyypit voivat aiheuttaa merkittäviä muutoksia luustolihaksen kuiduissa, vaikka ei ole mahdollista varmuudella vahvistaa, mikä oli samojen henkilöiden aikaisempi geneettinen rakenne. Tämä prosessi "voitaisiin" sallia makrojoukkoon II kuuluvien kuitujen erikoistumiskapasiteetilla tai jopa vain osalla.
On mahdollista, että tyypin IIx kuitujen hapetuskapasiteetti on parantunut korkean intensiteetin kestävyysharjoittelun jälkeen, mikä johtaa niihin tasolle, jossa ne kykenevät täyttämään oksidatiivisen aineenvaihdunnan yhtä tehokkaasti kuin kuidut I kouluttamattomilla henkilöillä.
Tämä määräytyy mitokondrioiden koon ja lukumäärän kasvun ja niihin liittyvien muutosten mukaan, mutta ei kuidutyypin muutoksella..




















-propriet-nutrizionali.jpg)