Jos solu toimii anaerobisissa olosuhteissa, se tuottaa energiaa muuttamalla glukoosin laktaatiksi ja hävittää sen Cori -syklin aikana; jos happea on saatavilla (siis lepotilassa), yli 90% glukoosista kulutetaan aerobisesti ja vain loput 10%, anaerobisesti. Kun tarvitaan enemmän ATP: tä kuin mitä aerobinen reitti pystyy tarjoamaan (esimerkiksi kun lihakset ovat rasittuneita), lisävarusteena on anaerobinen aineenvaihdunta (olemme hapen puutteessa: hengenahdistus, väsymys) hengitys jne.): On tarpeen nopeuttaa tätä aineenvaihduntaa muuttamalla laktaatti (joka saadaan glykolyysistä) glukoosiksi glukoneogeneesin kautta.
Aerobinen aineenvaihdunta kehittyy mitokondrioissa.
Ensimmäinen aerobisessa aineenvaihdunnassa havaittu entsyymi on pyruvaattidehydrogenaasi; On tarkempaa sanoa, että pyruvaattidehydrogenaasi on entsyymikompleksi entsyymin sijasta, koska se on 48-60 proteiiniyksikön aggregaatti, jossa on kolme katalyyttistä kohtaa peräkkäin.
Pyruvaattidehydrogenaasi katalysoi seuraavan reaktion (redoksi):
Pyruvaatti + NAD + + CoA-SH → Asetyyli CoA + NADH + H + + CO2
CoA-SH on koentsyymi A: se on pantoteenihapon johdannainen; asetyylikoentsyymi A on tioesteri. Tämä on redoksiprosessi, koska ensimmäinen pyruvaatin hiili menee hapetusnumerosta kolme hapetusnumeroon neljä (se hapettui) ja toinen pyruvaatin hiili muuttuu hapetusnumerosta kaksi hapetusnumeroksi kolme (se hapettui). Sitten pyruvaatti hapetetaan (se menettää yhteensä kaksi elektronia) ja NAD pelkistyy.
Kuten mainittiin, pyruvaatidehydrogenaasilla on kolmenlaisia entsymaattisia aktiivisuuksia, joista kukin tukee omaa katalyyttistä kofaktoriaan:
- tiamiinipyrofosfaatti (se on B1 -vitamiinin johdannainen); se on aktiivinen deprotonoituneessa muodossa: karbanioni muodostuu.
- lipoamidi (se on lipoiinihapon johdannainen); se sisältää erittäin reaktiivisen disulfidisillan.
- flaviini -adeniinidinukleotidi (se on B2 -vitamiinin johdannainen); se on nukleotidi, jolla on redox -ominaisuuksia: sen redoksikeskus koostuu flaviinista.
Eukaryoottisoluissa aerobinen aineenvaihdunta tapahtuu solun erikoisorganelleissa, jotka ovat mitokondrioita; bakteereissa glukoosin ja muiden lajien aineenvaihdunta tapahtuu solussa, mutta ei ole erikoistuneita organelleja.
Kun pyruvaatti pääsee mitokondrioihin, se altistuu pyruvaattikarboksylaasin vaikutukselle, jos glukooneogeneesi on suoritettava (lähtöaineen rekonstruoimiseksi), tai se voidaan altistaa pyruvaatidehydrogenaasille, jos se on tarpeen energian tuottamiseksi: "Asetyylikoentsyymi A, joka muodostuu aerobisella aineenvaihdunnalla, stimuloi pyruvaattikarboksylaasin toimintaa, joten se edistää glukoneogeneesiä ja vähentää pyruvaattidehydrogenaasin toimintaa.
Katsotaan nyt, miten pyruvaattidehydrogenaasi toimii; ensinnäkin pyruvaatti dekarboksyloituu tiamiinipyrofosfaatin vaikutuksesta.
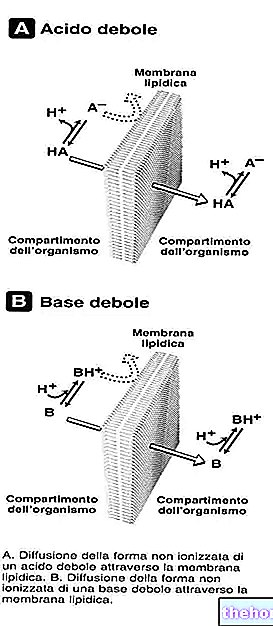
Happama ympäristö voi estää aerobista aineenvaihduntaa, koska tiamiinipyrofosfaatin anioninen muoto on aktiivinen, joka protonoituisi happamassa pH: ssa ja dekarboksylaatiota ei tapahtuisi.
Dekarboksylointi on vaikea reaktio, koska hiili-hiiliside on katkaistava; tässä tapauksessa reaktiota suosii termodynaamisesti se, että reaktiovälituote (hydroksietyylitiamiinipyrofosfaatti) antaa resonanssin (molekyylin p-elektronit delokalisoidaan): hydroksietyylitiamiinipyrofosfaatti esiintyy kolmessa mahdollisessa muodossa (resonanssi) ja tämä tekee siitä melko vakaan. Lisäksi anionisessa muodossa oleva hydroksietyylitiamiinipyrofosfaatti säilyy riittävän kauan voidakseen olla vuorovaikutuksessa lipoamidin disulfidisillan (pyruvaattidehydrogenaasin toinen katalyyttinen kofaktori) kanssa; disulfidisilta on värähtelevä varsi (se sijaitsee pitkän joustavan ketjun pää) ja voi siirtyä katalyyttikohdasta toiseen entsyymikompleksissa.
Sitten lipoamidi sitoo disulfidisillan kautta hydroksietyylitiamiinipyrofosfaattia: saadaan asetyylilipoamidia.Tämä on transetetylointireaktion ensimmäinen vaihe, jota katalysoi pyruvaattidehydrogenaasikompleksin ensimmäinen entsyymi; tässä vaiheessa se katkaisi siteen hydroksyyliryhmä ja tiamiinipyrofosfaatti, jotka palasivat alkuperäiseen muotoonsa: tapahtui redoksireaktio, jossa disulfidisilta toimi hapettimena (kaksi rikkiatomia pelkistyi) kohti hydroksyyliryhmää, jonka se hapetti asetyyliksi.
Tämän vaiheen jälkeen lipoamidin värähtelevä käsivarsi liikkuu ja lähestyy toista pyruvaattidehydrogenaasin entsyymiä, joka suorittaa todellisen transasetylaasiaktiivisuuden kantamalla asetyyliryhmän mukanaan: tapahtuu toisen entsyymin katalysoiman transasetylointireaktion toinen vaihe; Tällä tavalla olemme saaneet asetyylikoentsyymi A. Nyt on tarpeen palauttaa lipoamidi, joka on pelkistetyssä muodossa: kolmas pyruvaatidehydrogenaasin entsyymi puuttuu asiaan, mikä hapettaa lipoamidin uudelleen ja siirtää sen elektrodit FAD: iin, joka pelkistetään FADH2: ksi. FAD / FADH2 voi toimia redoksiparina kahdessa erillisessä monoelektronisessa vaiheessa tai yhdessä bielektronisessa vaiheessa.
FADH2 antaa välittömästi elektroninsa NAD +: lle, jolloin saadaan FAD ja NADH + H +.
Asetyylikoentsyymi A, joka on saatu kuvatulla tavalla, on Krebs -syklin (tai trikarboksyylihapposyklin) lähtöaine.


.jpg)