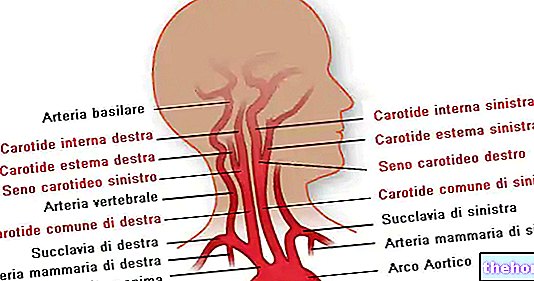
Krebs -sykliä kutsutaan myös trikarboksyylihapposykliksi ja siinä käytetään asetyylikezimiimi A: n lähtömetaboliittia, joka saadaan pyruvaattidehydrogenaasin vaikutuksesta glykolyysin tuottamaan pyruvaattiin.
ATP ja pelkistysteho saadaan krebs -syklistä; pelkistävä voima lähetetään hengitysketjuun, jossa NADH ja FADH2 hapetetaan vastaavasti NAD +: ksi ja FAD: ksi: pelkistävä teho siirtyy hengitysketjua pitkin kytkentäjärjestelmiin, joista tuotetaan lisää ATP: tä.
Krebs -sykli on hermokeskus paitsi glukoosin aineenvaihdunnalle myös rasvahappojen ja aminohappojen aineenvaihdunnalle, itse asiassa asetyylikoentsyymi A: ksi muuttuva pyruvaatti ei johdu pelkästään glukoosin hajoamisesta: se saadaan esimerkiksi alaniinin (aminohapon) transaminoinnista.
Noin 80% Krebs -sykliin osallistuvasta asetyylikoentsyymistä A tulee rasvahappojen aineenvaihdunnasta.
Asetyylikoentsyymi A on tioesteri, joten sen energiapitoisuus on korkea sitraattisyntaasi uuden hiili-hiilisidoksen muodostamiseksi; sitraattisyntaasi on krebs -syklin ensimmäinen entsyymi.
Asetyylikoentsyymi A: n metyylihiili vapauttaa vapaaehtoisesti (tautomerismin avulla) protonin (tulee karboanioniksi) ja hyökkää oksaloasetaatin karbonyylihiileen: muodostuu korkean energian pitoisuus tioesteri (sitriilikoentsyymi A), josta sitraatti saadaan hydrolyysillä ja koentsyymi A uudistetaan. Sitraattisyntaasia moduloivat negatiivisesti tuote, ts. sitraatti ja ATP: jos sitraatti kertyy, tämä tarkoittaa, että tämä vaihe on nopeampi kuin muut, joten sen on hidastuttava (sitraatti on negatiivinen modulaattori).

ATP vaikuttaa myös sitraattisyntaasin toimintaan, koska pelkistävä teho saadaan krebs -syklistä, joka sitten lähetetään hengitysketjuun, josta ATP tuotetaan; jos ATP kertyy, se tarkoittaa, että tuotetaan enemmän kuin tarvitaan. Hidastamalla krebs-sykliä (sykli hidastuu, jos jokin sen vaiheista hidastuu) myös ATP: n tuotanto hidastuu: ATP: n negatiivinen modulaatio on palautemodulaatio (yhden lopputuotteen muodostuminen on moduloidaan säätämällä prosessin yhden vaiheen nopeutta).
Krebs -syklin toisessa vaiheessa sitraatti muuttuu entsyymin vaikutuksesta isositraatiksi akonitaasi; entsyymin nimi johtuu siitä, että sitraatti dehydratoidaan ensin muodostaen cis-akonitaattia ja sen jälkeen vesi palaa takaisin kiinnittymällä hiileen, joka on eri kuin se, johon se oli aiemmin sitoutunut. Isositraatti saadaan ilman, että substraatti poistuu katalyyttikohdasta; akonitaasi on stereospesifinen entsyymi: se tunnistaa sitraatin kolme karboksyylikeskusta ja saa sitraatin pysymään sitoutuneena entsyymiin niin, että "veden poistuminen ja sisäänpääsy" cis-aconitate-välituotteen läpi.
Krebs -syklin kolmannessa vaiheessa on ensimmäinen energiahuomautus, koska hiilidioksidina eliminoitu hiili häviää. Entsyymi, joka katalysoi tätä vaihetta, on isositraattidehydrogenaasi; alustalle suoritetaan ensinnäkin dehydraus: NAD + saa pelkistävää voimaa ja muodostuu oksalosukkinaattia (se on meripihkahapon oksaalijohdannainen).
Entsyymi -isositraattidehydrogenaasilla on kaksi modulaatiokohtaa: positiivinen modulaatio ADP: n vuoksi ja negatiivinen modulaatio ATP: n vuoksi. Päivittäin kulutettavan ATP: n määrä on erittäin korkea: ATP tuottaa hydrolyysinsä "ADP: n ja kaiken" ortofosfaatin vapauttaman energian.
Nukleosidien (typpipitoinen emäs plus sokeri) ja nukleotidien (nuklosidi plus fosfaatti) kokonaispitoisuus elimistössä on lähes vakio: sanoa siis, että c "on paljon ATP: tä tai vähän ADP: tä (tai päinvastoin, paljon ADP: tä) ja pieni ATP) on sama asia; ADP on synonyymi energiantarpeelle ja on siksi positiivinen modulaattori, kun taas ATP on oire energian saatavuudesta ja on siksi negatiivinen modulaattori.
JATKA: Toinen osa "













.jpg)