Katso myös: ketogeeninen ruokavalio; diabeettinen ketoasidoosi.
Yleisyys
Aiemmin luultiin, että ketonikappaleet johtuvat liiallisesta aineenvaihdunnasta, joka johtuu liiallisen rasvan nauttimisesta tai diabeteksesta. Ketonikehot puolestaan ovat luonnollisesti kehomme tuottamia: aivot sopeutuvat käyttämään näitä metaboliitteja olosuhteissa pitkäaikainen paasto (diabeetikoilla ketonikappaleet korvaavat glukoosin aineenvaihdunnan).
Mitkä ovat ketonikappaleet?
Ketonirungot ovat lipidijohdannaisia (ne ovat peräisin lipidien aineenvaihdunnasta, lähes yksinomaan maksasta), mutta niillä on ominaisuuksia, jotka tekevät niistä samanlaisia kuin sokerit:
- Suuri syöttönopeus;
- Nopea käyttää.
Jopa jotkut aminohapot, erityisesti aineenvaihduntaolosuhteet, voivat tuottaa ketonikappaleita (leusiini, lysiini, fenyylialaniini, isoleusiini, tryptofaani ja tyrosiini).
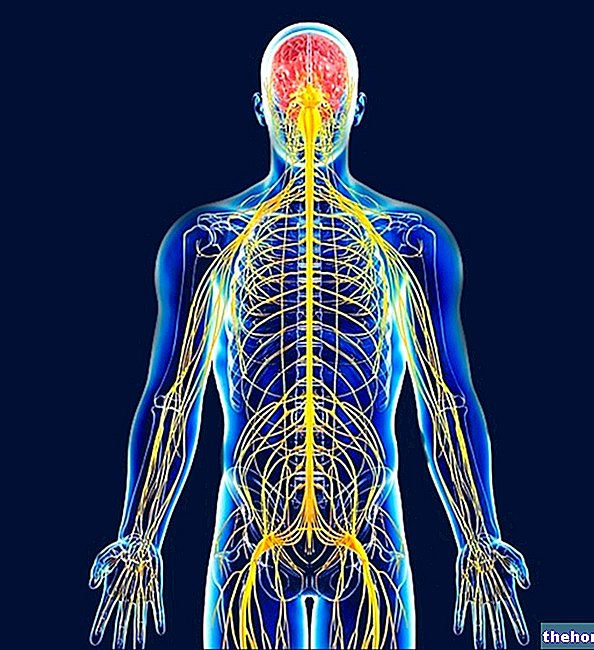
Biologinen rooli
- Ketonirungot ovat kooltaan pieniä, joten ne kuljetetaan erittäin nopeasti (paljon enemmän kuin rasvahapot, jotka toisaalta tarvitsevat kuljetusproteiineja, kuten albumiinia);
- ketonikappaleita käyttävät lähes yksinomaan lihakset ja perifeeriset kudokset, mutta myös sydän (20-30% sen käyttämästä energiasta tulee ketonikappaleista) ja aivot (pitkäaikaisen paaston tapauksessa).
Synteesi
Ketonirunkoja syntetisoi asetyylikoentsyymi A, joka on peräisin rasvahappojen aineenvaihdunnasta.
Entsyymi, joka katalysoi ensimmäisen vaiheen, on Β-ketotiolaasi, joka käyttää asetyylikoentsyymi A: n rikkiä produce-ketoasyylikoentsyymi A: n tuottamiseen (se on päinvastainen reaktio kuin rasvahappojen Β-hapetuksessa); tämä reaktio ei ole spontaani, vaan sitä ohjaa seuraava reaktio , katalysoitu "hydroksimetyyliglutaryylikoentsyymi A -syntaasi ja johon liittyy toisen asetyylikoentsyymi A: n kiinnittäminen, jolloin saadaan 3-hydroksi-3-metyyliglutaryylikoentsyymi A.
Tämän jälkeen puuttuu lyyttinen entsyymi, joka muuttaa 3-hydroksi-3-metyyliglutaryylikoentsyymi A etikka-asetaatiksi, joka on ketonirunko. Etikka -asetaatti voidaan lähettää perifeerisiin kudoksiin tai entsyymin vaikutuksesta hydroksibutyraattidehydrogenaasi, muunnetaan 3-Β-hydroksibutyraatiksi. Jos etikka -asetaatti on erittäin suuri pitoisuus, se voi myös spontaanisti dekarboksyloitua asetoniksi.
Asetoni, etikka-asetaatti ja 3-Β-hydroksibutyraatti ovat kolme harkitsemamme ketonikappaletta; asetoni on jätetuotetta, joka muodostuu satunnaisesti ketonikappaleiden reitille ja poistuu uloshengityksen ja haihtumisen kautta.
Käyttö perifeerisissä kudoksissa
Maksassa tuotetut ketonikappaleet lähetetään perifeerisiin kudoksiin.
Katsotaan nyt, mitä tapahtuu, kun etikka-asetaatti ja 3-Β-hydroksibutyraatti saavuttavat perifeeriset kudokset. Etikka-asetaatti on Β-ketohappo, joten aktivoituna sitä voidaan käyttää Β-hapetusprosessissa asetyylikoentsyymi A: siksi on tarpeen muuttaa Β-ketohappo Β-ketoasyylikoentsyymi A: ksi.
Kun etikka -asetaatti saapuu perifeerisen kudoksen solun mitokondrioihin, se altistuu entsyymin vaikutukselle sukkinyylikoentsyymi A -transferaasi: tämän entsyymin kautta etikka -asetaatti reagoi sukkinyylikoentsyymi A: n kanssa (tulee Krebs -syklistä) ja saadaan sukkinaatti ja etikka -asetyylikoentsyymi A.
Hyödyntämällä sukkinyylikoentsyymi A: ta etikka -asetaatin aktivoimiseksi hyppäämme Krebs -sykliin, vaiheeseen, joka tuottaa GTP: n - tämä on energiaprosessi, jonka solu on valmis maksamaan asetyylietikka -koentsyymi A: n saamiseksi ; jälkimmäinen menee sitten toiminnan alle Β-keto-tiolaasi (Β-hapetusentsyymi) tuottaakseen kaksi asetyylikoentsyymi A -molekyyliä, jotka lähetetään krebs-sykliin.
Jos 3-Β-hydroksibutyraattia lähetetään perifeerisiin kudoksiin, jälkimmäinen muuttuu mitokondrion sisällä etikka-asetoniksi Β-hydroksibutyraattidehydrogenaasientsyymin vaikutuksesta, jolloin muodostuu NADH, joka vastaa noin 2,5 ATP: tä; valmistettu etikka -asetaatti seuraa edellä kuvattua tietä.
Perifeerisen kudoksen solu imee enemmän energiaa 3-Β-hydroksibutyraatista kuin etikka-asetaatista, mutta yhden tai toisen kuljetus perifeerisiin kudoksiin riippuu maksan energian saatavuudesta.
C "on vähäinen määrä metaboloituneita rasvahappoja, jotka sisältyvät peroksisomeihin eivätkä mitokondrioihin; peroksisomit ovat mitokondrioita pienempiä organelleja ja runsaasti metalli-ioneja ja peroksidaasientsyymejä. Peroksidaasientsyymit käyttävät vetyperoksidia edistämään redoksiprosesseja, joten peroksisomeissa on entsymaattinen järjestelmä, joka pystyy tuottamaan vetyperoksidia.
Peroksisomien Β-hapetuksessa "asyylikoentsyymi A saadaan aikaan"asyylikoentsyymi A -oksidaasi (Toisaalta mitokondrioissa asyylikoentsyymi A -dehydrogenaasi -entsyymi toimi.) Myös tässä tapauksessa muodostuu trans -2,3 -enoyylikoentsyymi A, joka käy läpi bifunktionaalisen entsyymin (se suorittaa saman tehtävän kuin mitokondrioissa "enoyylikoentsyymi A -hydrataasi ja L-Β-hydroksiasyylikoentsyymi A -dehydrogenaasi") ja muuttuu siten Β-ketoasyylikoentsyymi A: ksi. ja saadaan asyylikoentsyymi A, jossa hiilipitoinen luuranko on vähennetty kahdella yksiköllä verrattuna lähtöyksikköön, joka palaa verenkiertoon.
















-perch-e-quando-si-misura.jpg)