Elävien olentojen syklinen jatkuvuus löytää sukupolvien väliset yhteydet lisääntymisilmiöissä.
Jäljentäminen tapahtuu evoluutiomittakaavan eri tasoilla, kasvi- ja eläinvaltakunnan erilaisissa seurauksissa, eri elävissä lajeissa, niin monilla mekanismeilla, jotka yksinään oikeuttavat koko tutkielman.
Ensimmäisessä lisääntymisilmiöiden luokittelussa on erotettava yksisoluiset monisoluisista organismeista, koska vain ensimmäisessä solujen jakautuminen on sama kuin lisääntyminen.
Monisoluisessa lisääntyminen voi olla agamista tai seksuaalista (tai gammalaista).
Agaminen lisääntyminen, joka on suhteellisen harvinaista, perustuu mitoosimekanismiin, joten lajin vaihtelevuus on uskottu mutaatioiden toistuvalle esiintymiselle.
On myös erilaisia mekanismeja, kuten strobilisaatio, itiöinti jne., Kun taas kasveista löydämme maataloudessa hyvin tunnettuja uudistavia muotoja (pistokkaat, kerrostumat jne.).
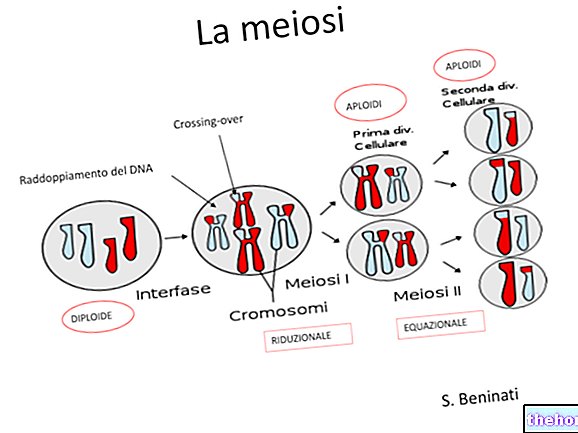
Yleisin lisääntymismekanismi korkeammissa muodoissa on kuitenkin seksuaalinen mekanismi, joka vastaa meioosin esiintymistä, sukusolujen muodostumista ja niiden sulautumista tsygoottiin (hedelmöitys).
Alkukantaisissa lajeissa sukusolut eivät ole morfologisesti eriytyneet: tässä tapauksessa puhumme isogametiasta. Kuitenkin aina on kaksi sukusolujen sarjaa, jotka on merkitty symboleilla (+) ja (-), ja hedelmöitys voi tapahtua vain "vastakkaisen merkin sukusolujen välisestä kohtaamisesta: siksi on olemassa biologinen ero, jota ei ole vielä ilmaistu morfologisesti.
Kehitysasteikon edetessä ilmenee morfologinen ja toiminnallinen erilaistuminen, jossa naaraspuolisten sukusolujen tyyppi, jolla on yleensä runsaasti vara -ainetta (deutoplasma tai vasikka, joka varmistaa alkion kehityksen, kunnes se voi olla aineenvaihduntaan riippumaton) ja eräänlainen urospuoliset sukusolut, joilla on liikkuvuutta päästä naaraspuolisiin.
Meíoosin ja tsygootin välillä voi kulua sarja haploidisolusukupolvia, aivan kuten tsygootin ja meioosin välillä voi kulua sarja diploidisolusukupolvia, jolloin eri sukupolvien lajit ilmaistaan sukupolvien vaihtelulla.
Diplonte -lajille (jolla on diploidinen organismi) on ominaista gametinen meioosi: meioosi tuottaa suoraan sukusoluja, jotka yhdistämällä välittömästi palauttavat diploidisen tilan. Tämä on yleinen tapaus metatsooilla, mukaan lukien ihminen.
Gametogeneesi
Kun otetaan huomioon lisääntyminen gametisella meioosilla, sellaisena kuin se tapahtuu ihmisellä, yritetään selventää, miten meioosi sopii gametogeneesiin (sukusolujen muodostuminen).
Alkionkehityksessä, miesten ja naisten gametogeneesissä (kutsutaan spermatogeneesiksi ja oogeneesiksi), varhainen erilaistuminen solujen välillä, jotka on tarkoitettu muodostamaan keho (somaattinen linja), ja solujen välillä, jotka on tarkoitettu tuottamaan sukusoluja (ituradat). Sukusolun alkusoluja kutsutaan protogoneiksi. Sukupuolielinten erilaistumisella miehen tai naisen mielessä sukusolut erilaistuvat vastaavasti spermatogoniaksi ja ovogoniaksi.
Tarkasteltaessa spermatogeneesiä näemme, että spermatogoniassa on sarja solusukupolvia, jotka jatkuvat koko elämän ajan. Vain osa tällä tavoin jatkuvasti tuotetusta spermatogoniasta eroaa normaalista mitoottisesta syklistä ja sen sijaan alkaa meioottinen.
Sukusolua, jossa meioosi alkaa (reduplikaatio ja sitten ensimmäinen jakautuminen), kutsutaan ensimmäisen asteen spermatosyytiksi; sen jakautumisesta syntyy kaksi toisen kertaluvun siittiöitä, joista toisen jakautumisen yhteydessä syntyy yhteensä neljä siittiöitä.
Voimme siirtyä kromosomaalisen kitin pienentämisestä 4n ensimmäisen asteen spermatosyyteistä (reduplikaation jälkeen on neljä kromatiilia kutakin homologiparia varten) toisen asteen spermatosyyttien 2n: een ja spermatidien n: ään, kuten jo on tutkittu Meioosi, josta päätellään.
Naisen gametogeneesissä (tai oogeneesissä) on useita eroja. Ensinnäkin valmistettavien sukusolujen määrä on paljon pienempi. On arvioitu, että noin 5 x 105 ovogonia valmistetaan ihmislajin naarasrauhasissa; Näistä vain noin 400 on kiinnostunut follikkelien kypsymisestä ja sen jälkeisestä hukasta syklissä, joka yleensä vaikuttaa vain yhteen follikkeliin kuukaudessa noin 35 vuoden hedelmällisen ajan.
Molemmilla sukupuolilla valmistettujen sukusolujen eri määrä vastaa jo mainittua toimintojen ja käyttäytymisen eroa: siittiöt ovat pieniä, liikkuvia ja lukuisia, kun otetaan huomioon tarve etsiä muna ja pieni todennäköisyys löytää se; munat ovat suuret, inertit ja harvat suhteessa tehtävään taata alkio varantoaineelle ja sisäisen lannoituksen tarjoamalle suojalle (luonnollisesti, erityisesti ulkoisen lannoituksen yhteydessä, myös munasolujen on oltava enemmän).
Tarve varustaa sukusoluja vara-aineella vastaa "oogeneesissä" meioosin pysäytysvaiheen läsnäoloa, jonka aikana kromosomit ovat osittain despiralisoituneet. Sitten havaitsemme niin sanottuja "höyhenkromosomeja", joissa sarja extroversioista tunnistaa piirteet, joissa deutoplasman synteesistä vastaavat geenit ovat lamaantuneet.
Naaraspuolisten sukusolujen vaatima pienempi määrä vastaa myös sitä tosiasiaa, että meioosin tuottamista neljästä haploidisolusta vain yksi saa koko vara -aineen ja siitä tulee sukusolu, kun taas kolme muuta (polosyytit tai polaariset kappaleet), jotka sisältävät vain kromosomimateriaalista, ei voi synnyttää zygootteja ja alkioita, ja niiden on määrä taantua.
Lannoitus
Lannoitus eli uros- ja naaraspuolisten sukusolujen kohtaaminen voidaan suorittaa hyvin eri tavoin.Eläinkunnassa havaitsemme siirtymisen ulkoisesta hedelmöityksestä (sukusolut, jotka ovat alttiita ympäristöriskeille ja siksi välttämättä erittäin lukuisia molemmilla sukupuolilla) sisäiseen hedelmöitymiseen, johon vanhempien hoito liittyy edelleen nisäkkäiden aineenvaihduntasuhteeseen äidin ja sikiön välillä .
Hedelmöityksen on tapahduttava vastakkaista sukupuolta olevien sukusolujen kohtaamisen jälkeen, mikä takaa kaksi ehtoa: spesifisyyden ja ainutlaatuisuuden. Toisin sanoen on varmistettava, että siittiöt ovat samaa lajia kuin muna ja että kun ensimmäinen tulee, muut eivät pääse sisään.
Spesifisyyden takaavat akrosomin ja munasolun pinnan biokemialliset ominaisuudet. Itse asiassa puhutaan "hedelmöitysaineiden" ja "antifertilisiinien" välisistä reaktioista, joiden spesifisyys on verrattavissa entsyymien ja substraatin kohtaamiseen.
Lannoituksen ainutlaatuisuus taataan munasolun pintarakenteen muutoksella ("kortikaalinen reaktio"), joka alkaa heti ensimmäisen spesifisen lannoite / hedelmöitysvastaisen reaktion hetkellä; tämän reaktion jälkeen munasolun kalvo muuttuu niin, että muut siihen saapuneet siittiöt eivät enää pysty aloittamaan spesifistä lannoitusreaktiota.
Hedelmöityksen jälkeen siittiöiden pyrstö pysyy munasolun ulkopuolella, kun taas kromosomimateriaali tunkeutuu siihen.Tämä, nimeltään "urospuolinen pronukleus", liittyy munasolun "naaraspuoliseen pronukleukseen" muodostaen tsygootin diploidisen ytimen.